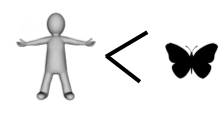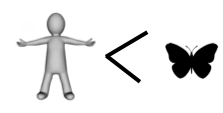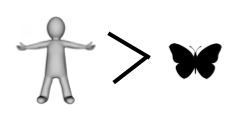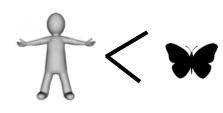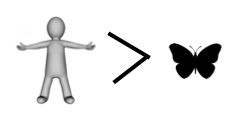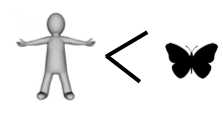I am inaugurating this blog series with a virtual visit to Nectandra Cloud Forest, to set the stage for an introduction to an ecosystem that’s is rapidly shrinking. Thirty years ago, when the scientists first described what is a cloud forest, they estimated around 11% of the world tropical rain forests to be cloud forests — forests that are immersed in cloud most of the year. Today, it is estimated that only 1.5% is left and disappearing. At today’s rate of climate change, most of it will be gone before we know it. Rather than lament its impending loss, I would like to use the time and opportunity to celebrate this Garden of Eden and to share with you our experiences, the highlights and the sense of wonderment living in such an environment.
Imagine yourself dangling from a parachute, floating straight down on Nectandra forest looking down through your feet at 150 m (500ft) in the air.
You are about to land on what appears to be an immense head of broccoli where each tree constitutes a floret. But unlike the florets, the trees in the canopy are not clones. No two look alike because they are likely to be of different species. They come in all shades of green, each with a different texture and coarseness.
In the image above, the tightly packed tree canopy shows hardly any gaps between trees and even less between foliage. Tree branches are not visible. In fact, the degree of gaps in the forest can be taken as an inverse measure of the density of the forest. The denser the trees, the less gap areas in the canopy. Scientific computer software can count the black or gap pixels per unit screen area as a way to measure and monitor forest growth in drone images .
Drop down another 50 m and look ahead to get a different view.
The trees appear now more varying in shapes, in heights and no longer pure green. Under the oblique light, some of the trees definitely take on a brownish hue. Leaves changing color? Dying trees? Look closer and you will note that the brownish green is not foliage, but thick mats of plants on the branches and trunks of trees. The brownish growth are hundreds of intergrowing species of the liverworts and mosses — the tiny iconic plants of the cloud forest known collectively as the bryophytes.
A startling different view emerges as you descend to about 15m from the ground. You are now just below the upper canopy, looking through the tangles of the branches. The presence of the bryophytes is overwhelming.
The accumulating bryophytes and plant detritus provide the necessary nutrients and loamy support for other epiphytic flora (plants that grow on other plants). Add the steady, constant misting during the day all year to the mix, plants with aerial roots can now grow and thrive. Well known examples are the bromeliads (air plants), orchids, ferns, lianas, even begonias, to name just a few. Shown in the photo immediately below are impressive inflorescence of hanging orchids Oncidium sp. (see inset) on the left and an endemic Columnea macrophylla on the right (in the same family as african violets). Visitors on the ground are likely to miss this floral hanging garden because of the altitude.
From this height another striking, eye-catching group of cloud forest iconic trees are the tree ferns. The dozen species of tree ferns at Nectandra are beauties to behold, with their majestically large fronds (~1.5m) and great stature (up to 10-15 m).
Let’s drop down to the forest floor. Now look up. The view is typically through thick foliage of understory plants, and through epiphytes-laden branches. From this vantage point, nearly every surface is layered with bryophytes and epiphytes.
Look down, more bryophytes, on tree trunks, branches, leaves, rocks and soil…
Zoom in on the tree trunks, now the moss (below left) can be distinguished from the intermingling liverworts on the right, two representatives species among hundreds on the reserve.
We have come to the end of our vertical tour. On your future visits, I hope to introduce you to many more forest features and riveting fellow residents. Please check in for our next blog for more unconventional virtual tours of the Nectandra Cloud Forest.
…..a chance encounter, an unexpected discovery, a happy accident, a startling fluke, an unforeseen experience, a fortuitous involvement, an unintended contact, a sudden insight, and so many more . . .
Serendips challenge our mind’s readiness for discovery. They often give no warning, and do not advertise. Our ability to detect and experience them rests on our mind’s readiness and alertness.
Cigar-sized caterpillar of Automeris moth
Just as all irrepressible children with their memorable discoveries, we at Nectandra want to share the excitement of our numerous serendips with the family members, friends and anyone who share our love for Nature and her profoundly beautiful living creations.
Starting this August, we will post any unusual findings and observations on subjects of tropical nature in general, and of cloud forest in specific with readers of our blog serendips.net The articles will be short and entertaining (we hope) and definitely informational. Please subscribe to register your interest in becoming a Serendipper at Nectandra — the realm of furry plants and mossy animals .
Of the 300,000 estimated insects in Costa Rica, two groups of insects get my vote for celebrity status. They are the katydids (bush crickets) and phasmids (stick insects, the subject of the next blog). Found all over the world except in Antarctica, they are colorful, charismatic, and masters of disguise. Most of the species live in the tropics.
Katydids (Tettigoniidae)
Katydids appears similar to their grasshopper relatives, but with super long antennae many times its body length. They are tree-living omnivores and are the major nocturnal noise maker in the tropics. The song is a multisyllabic “ka ty did she did“)” mating call, generated by the males’ stridulation between the forewings. Ranging from 5 – 130mm in size, they feed on plants, insects, insect eggs and even small lizards. It is no coincidence that their coloration often matches parts of plants precisely, rendering them nearly invisible in their diurnal activities, but their tricks involve more than colors, as revealed in the examples below:
Fig 1A Katydid Copiphora rhinoceros (8cm). Katydids can move at lightning speed to catch its preys. Fig 1B The unihorn between its eyes is a defense mechanism against predators. So are its strong and sharp mandibles.
For comparison, grasshoppers are evolutionary older relatives of katydids (250 vs 23-66 million yrs). They have more powerful hind legs and shorter antennae than katydids (Fig 2). Grasshoppers can jump 20 times its body length. In human terms, that would be equivalent to the length of a football field. Mostly ground dwellers, they are strict vegetarians. Under certain conditions, they grow to enormous population density and form swarms. With their voracious appetite, they can become devastating crop pests. Grasshoppers also stridulate to attract potential mates. However, unlike katydids’ wing-to-wing grating, grasshoppers rub the hind legs against their wings.
Fig. 2 An unidentified grasshopper at Nectandra.
Life Cycle
During courtship, the male katydid sings and offers an unusual nuptial gift to the females. The gift consists of a proteinaceous, gelatinous gland attached to the sperm sac (Fig 3). Receptive females consumes the protein rich gift after mating. This nuptial meal increases, not the fertility (number of offspring) of the female, but the exclusivity of paternity and the fitness of the offspring.
Fig 3. White spermatophylax on a male katydid trying to attract the opposite sex at Nectandra.
Katydid life cycle start with the eggs deposited on twigs, Fig 4. The eggs are laid in regular overlapping rows.
Fig. 4 Katydid eggs on a branch.Photo credit. Seph. Wikicommons
Fig. 5 Two katydid juveniles (magnified in inserts) at different life stages on Oncidium orchid.
Fig 6 Wingless juvenile katydid.
Fig 7 Final molt of a katydid. Note the winged, mature katydid emerging from the wingless old molt.
The wingless hatchlings (Fig 5 & 6) have the appearance of the adults. They remain wingless through the 4-5 molts until they reach adulthood at the last molt Fig. 7.
Crypticity
The most remarkable katydid features are their cryptic leaf-shaped forewings with color, textures and shapes indistinguishable from their leafy hiding places. Their wings mimic the venation and coloration of green or dry foliage. Katydids can fly but prefer to stay motionless to avoid predation. Below are examples of several masters of disguise (Fig 8A – 8E).
Fig. 8A – 8E. Leaf katydids
Katydid wings mimics foliage not just in color, but in textures and astoundingly plant-like ribbing, complete with imitation chewed marks, cracking, and other leafy imperfections and discoloration. This pair of wings was recognized as insect wings (Fig 9) only because they were found attached to the remains of katydid and were exact mirror images of each other — an impossible floral occurrence !
Mostly green and brown in color, a rare recessive genetic mutation in katydids result in shocking pink katydids (Fig 9 A). Thus far, nothing is known about the survival (dis)advantages of the spectacular pink katydids.
Fig. 9A Pink katydid . The shocking pink color is the result of a recessive gene mutation.
Fig. 9B Lesser angled-wing katydid normal green variant.
One evening at home in Costa Rica three years ago, I received an uncharacteristically excited call from my neighbor. “You would never believe this. There is a wild oncilla in my yard! It’s hiding underneath one of the bromeliads! It’s alive but not looking well. Come quick if you want to see one!” I flew out the door in seconds, all fired up and just as excited as my neighbor to see the most elusive and smallest of the country’s six wild cats.
Fig. 1. Oncilla, commonly known as tiger cat, Leopardus tigrinus
I was completely smitten on first sight. It was no bigger than an average domestic cat, but what a spectacular coat of spots, including the belly! The small, delicate, winsome animal was semi-hidden under the vegetation. It made a tiny head motion when I carefully lifted the foliage, but otherwise remained silent and immobile. It was clearly moribund but had no sign of physical injury. We quickly took its photo, retreated, and observed it from a distance in the dark. The next morning, its body was found in exactly the same position. It had died sometime during the night. My neighbor buried it that same day. This was my one brief and sad introduction to the rare oncilla, my first in two decades.
Cat sightings at Nectandra are infrequent, but all six Costa Rican wild cats are present, known through casts of footprints (Fig 2), or camera trap images (Fig 3).
Fig 2. Plaster of paris cast of a jaguar footprint on Nectandra muddy trails.Fig 3. Puma captured by camera trap at Nectandra.
Our preserve is fronted by a sharp curve on a national highway. Occasionally, we saw injured animals hit by a car in front of our gate, as with the margay photographed (Fig 4 ).
Fig. 4 Margay killed by a car in front of the Nectandra Preserve. Note the long tail in relation to its hind legs, a distinguishing feature for margays.
By size, the six Costa Rican wild cats fall in three groups: the biggest are the jaguars and pumas (45 – 70 kg), ocelot and jaguarundi (11 – 13 kg) in the mid range, and the margay (2-5 kg) and oncilla the smallest (1.5 – 3 kg). I have seen two male jaguarundis in action, fighting on a tree after dark. They were impressively fierce, aggressive and making frightful noises. By size, coloration, the lack of spots, behavior, they were jaguarundis. I was certain they were not jaguars or pumas. Ocelot and margay (Fig 4), unfortunately, I have seen only as road kills in front of our gate. The smallest oncilla, known also as tiger cat, however, has been very elusive and rare. No one among our staff, whether on day or night duty, have seen one, dead or alive.
Given the small size of our sick cat, we excluded ocelot in the differential identification. But could it have been a margay? Margay has a longer tail (~1.5 X) in relation to its hind legs (Fig 4), whereas our cat’s tail was just slightly longer than its hind legs. Its coloration and spots were compatible with those described or seen in published photographs of tiger cats, we provisionally identified it as an oncilla. Armed with its photo and our brief observations made in the dark to aid in the identification, I went on a deep literature search for confirmation and more information.
It was obvious at once, from my reading, that among the American small spotted cats — the ocelot, margay and oncilla gave the experts the most difficulty. They shared overlapping geography, temporal distribution, and ecology. With slight variations, they overlap in sizes, are all spotted with rosettes and have similar coloration. In fact, expert mammalogists often could not tell them apart either, especially between the margays and oncillas. From late 1700’s onward to 2000’s, literally dozens of different specie names were attributed to one or both of the cats. Their taxonomy and phylogeny were unclear and remained in disarray until the advent of DNA technology.
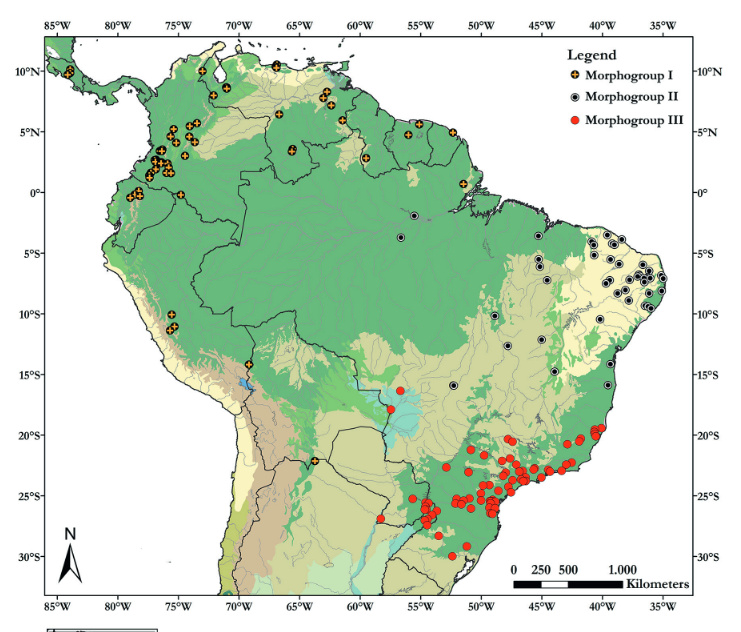
The low population density of oncillas was part of the problem. Sightings were spotty and infrequent. They are known from Costa Rica and Panama in the north, and in Columbia, Brazil, Bolivia in the south. Suitable specimens were few and far in between for comparative examinations. In one 2017 study, the authors were able to round up, from 23 museums in both Americas, only 250 skins and skull specimens to examine in detail. They analyzed the specimens’ associated geographic data, combined with careful skull measurements, survey of skin coloration, appearance of spots. Their results showed the distribution of the oncillas to be in 3 non-overlapping geographic groups (Fig 5).
Fig. 5 Geographic distribution of skull and skin specimens from 23 natural history museums in north and south America. Illustration from F Oliveira do Nascimiento and A Feijo, 2017. Taxonomic revision of the Tigrina Leopardus tigrinus. Papéis Avulsos de Zoologia http://dx.doi.org/10.11606/0031-1049.2017.57.19
Their pelts also showed discernible differences in coloration when compared side by side (Fig 6). The authors proposed three distinct morphologic groups I, II and III and were given names of I = L. tigrinus; group II = L. emiliae and group III = L. guttulus.
Fig. 6 Representative pelts from morpho group I (L.trigrinus), II (L. emiliea) and III (L. guttulus) according to authors F Oliveira do Nacimiento and A Feijo.
Our sick oncilla, therefore belongs presumptively in the most northern tiger cat group, L. tigrinus, according to proposed taxonomic division of 2017. But even before that year was over, there were indications that newer results from more discriminating DNA technology did not fully align with the conventional taxonomy. The oncilla phylogeny was more complicated.
We are now in 2023 and information on oncilla biology is still changing and being modified at a rapid rate. New species of tiger cats are being discovered, via DNA technology, in Panama and Brazil. Relationships among the spotted cats are further being clarified, their geneology delineated and phylogeny realigned, as telltale signs of ancient interbreeding among them (margay and ocelot, oncillas and Geoffroy’s cats) are being discovered.
Ironically, while the small spotted cats are rare to scientific studies, they are not rare to the illicit animal fur trade. Between 1976 and 1985, an astonishing 350,000 L. tigrinus pelts were traded! The illegal hunting and fur trading has since decreased due to conservation awareness and anti hunting laws, but it has sadly not stopped.
Bonus brain teaser: On a totally different aspect of observing cats, did you know that a kitten can be both a solid and liquid? Yes, a liquid. This realization won Marc-Antoine Fardin, a physicist at Paris Diderot University, the Ig Nobel Prize 2017. As in the photo below, the kitty (not an oncilla!) “flows” into, i.e. assumes the shape of the stein, just as beer would!
Imagine yourself leading a small group of adults and children through the thick cloud forest at Nectandra — all at once talking, walking and gently herding the children to stay on the trail. Your attention is split multiple ways — the subject matter, the ebullient children, spotting fauna and flora, where to land your feet and watching out for the coiled jumping pit viper that may become your next stepping stone. On two occasions, my feet came within 8 inches of the nearly invisible viperous bundle. I recoiled just in time in the first case and my companion yanked me sharply aside in the second. I don’t have a painful story to tell but the responsibility for guides to walk ahead became crystalline clear. Those seconds of terror also fixed my dread (and curiosity) of viper fangs.
Venomous distractions
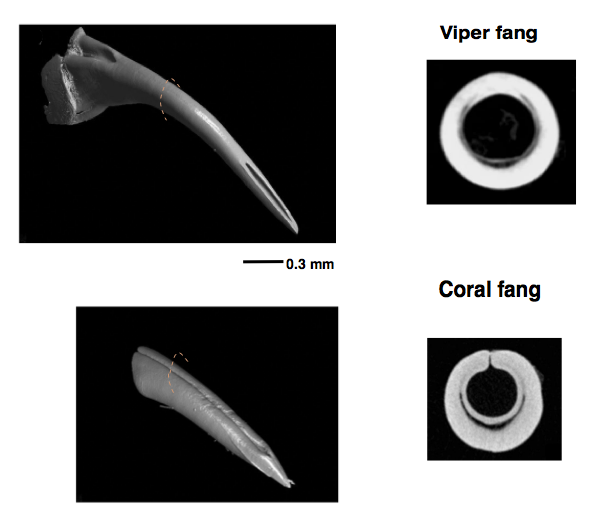
We all know about the danger of poisonous snake bites. We also know something about their venom and the availability of effective treatment using tailored antivenins. At Nectandra, elevation (1100 m, 3600 ft), pit vipers are more prominent than their better known cousin the fer-de-lance. The well camouflaged endemic jumping pit vipers (Fig 1 left) score the most bites requiring hospitalization in our area. The colorful, more timid coral snakes (Fig 1 right) have shorter fangs which may be loaded with neural venom, potent enough to be a source of worry for field workers and homeowners (I have caught several inside my house). Fortunately, our nearest hospital in San Ramon (15km) stocks antivenins to both snake groups.
While snake bites are professional hazards in the tropical forest, they are fortunately infrequent. Nevertheless, they are psychologically terrifying and medically serious. At our local hospital, the antivenin treatment requires two steps. If antivenin therapy is indicated, the first important step is to match the snake to the specific antivenin. The snake identity is determined from photos (thanks to cell phones), descriptions, physical specimens or from symptoms/appearance of the physical bites. If the snake identification is not possible (a very infrequent occurrence in Costa Rica), polyvalent antivenin to both the vipers and coral snakes is administered. In the second step, the patient is first infused with 1/10 of the final dose. If there is no detectable hypersensitivity to the associated horse serum, the infusion continues with the rest of of antiserum diluted in ≥40ml of saline. The whole procedure takes about 4-5 hours in the emergency room, followed by 3 days of observation.
Nationally, most (85%) of 500 annual snake bites are from the fer-de-lance (Bothrops asper). Coral snakes contribute ~2% of the bites. Even rarer are the bites from the jumping pit viper, an endemic snake found in mountainous, mid elevation region such as Nectandra. Unfortunately with a warming climate, the fer-de-lance has moved uphill and is now present at our cloud forest.
Why do snakes bite?
Snakes are able to carefully control the injection of venom. When threatened, 50% of the viper bites are “dry” bites with no or very small amount of the precious toxins injected, whereas most of the bites to kill prey involve venom. Hence, viper venoms are more for food and less for defense. In contrast, 85% of coral snake bites involve envenomation.
Anatomy of the jaws and fangs
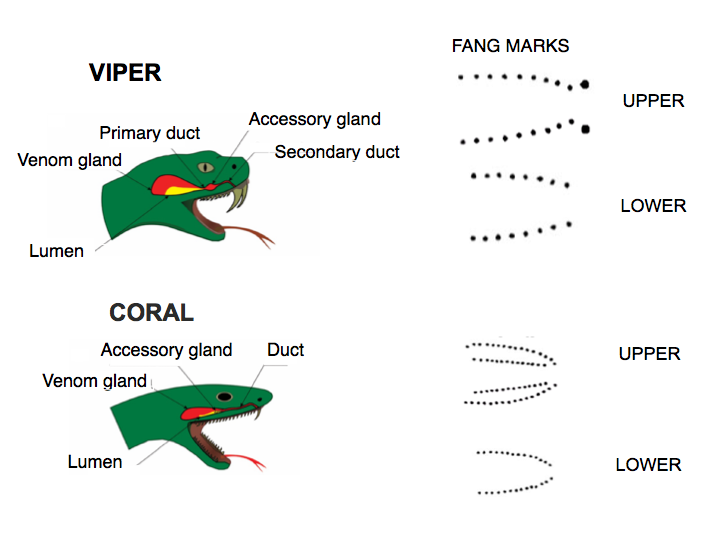
Snakes swallow prey whole. Mechanically, it is not an obvious nor trivial task for them to bite/ingest large animals many times relative to its body size. First, let’s look at the jaw anatomy. Their specialized mandibles are not attached to each other, but to stretchy ligaments allowing their jaws to open very wide to accommodate very large prey. In vipers, the mouth expands, the two long fangs (Fig 2 top), which are normally folded inside the mouth, swing out (like a blade from a switch knife) and sink in to grab the struggling prey. The fang puncture marks and bite-associated physical symptoms are hallmarks of viper bites. Rows of backward-pointing, smaller teeth help the snake grab and pull in the prey. In comparison to the viper, the coral snake has smaller, shorter, fixed fangs, and multiple rows of upper teeth. In spite of their teeth, snakes do not chew their food. Vipers depend entirely on the fast acting digestive enzymes in their venoms to feed. In encounters with humans, viper bites are sharp and brief. Strike and release. In comparison, the less aggressive coral snakes may be more reluctant to bite, but when they do, they are slow to let go.
Fig 2. Illustration adapted from Dehghani, Sedaghat & Fathi, Int Arch Health Sci, 2005
How is the venom delivered?
The venom is injected to paralyze, hemorrhage and break down tissues in prey. The snakes’ poison glands located behind their eyes are counterparts of the human’s salivary glands. The venom is specialized saliva containing a large cocktail of proteinaceous (> 50 enzymes) zootoxins. The venom is released from the system of glands through ducts under muscular pressure. It is injected much like liquid in a hypodermic needle and delivered through its hollow fangs under pressure of up to 30 psi (as in car tires). The size of the glands/ducts vary greatly, ranging from a few millimeters in length up to 1/4 of its length in the (old world) blue coral snake.
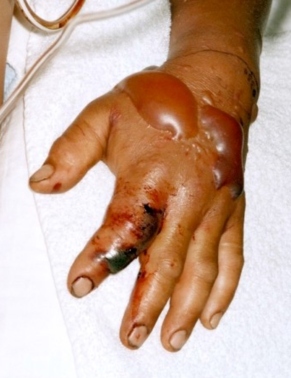
The vipers’ venom action in humans is rapid and intense. They may cause a combination of pain, welling, numbness, bleeding, bruising and blisters at the wound site (Fig 3). In comparison, coral venoms are neurotoxins and cardiotoxins . The smaller coral fang marks often are harder to see, painless with no swelling. The neurologic symptoms (drowsiness, nausea, headache, difficult breathing and paralysis) take longer to develop (up to 12 hours) but are then precipitous.
Fig 3. Photo from National Occupational Safety & Health, CDC
The best of fangs
The hollow, specialized teeth which we call fangs of the vipers (40 million yrs) and coral snakes (20-40 million yrs) evolved independently. As limbless animals, the snakes’ survival depend on their fangs, for both food and defense. The snakes’ successful radiation throughout the world attest to their fangs’ efficiency. Not only are snake venoms customized to their prey, but so are their fangs.
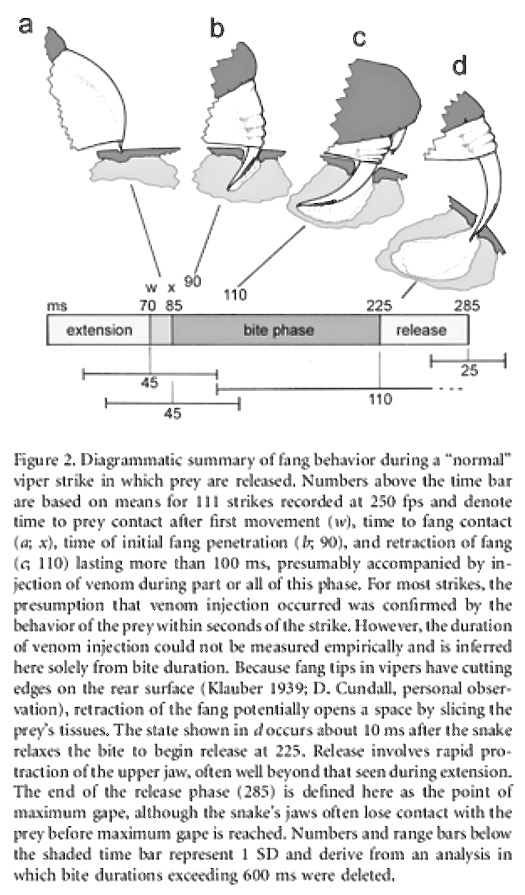
The fangs first task is to be able to penetrate the prey. The coral snake’s shorter and fixed fangs restrict the prey size. They must be able to bite through skins of reptiles, amphibians, insects and small mammals (e.g. scales, keratinized carapaces and furry skin etc). In contrast, vipers’ food may include all the above plus a wide range of large mammals and birds, varying in size, shape, hardness and textures. The complex mechanics of viper bites are being studied by a number of methods. For example, through time-lapse photography, the sequence of a viper bite when broken down into four steps, took place in an average of 285 milliseconds (about 3.5 bites per second) — during which time a small hole of the prey’s tissue was sliced open and the venom injected (Fig 4).
Figure 4. from David Cundall, Physiol Bioch Zoology Vol 82, 2009 https://www.jstor.org/stable/10.1086/594380
Newer technology such as microCT imaging reveals at high resolution the internal morphology and architecture of the fine bones and dentition structures in snakes (Fig 5).
Fig. 5. MicroCT scan of an adder’s head. Photo by L. Brian Stauffer, specimen FMNH11006 of the Field Museum of Natural History
Fig. 6 MicroCT scanning of viper fang (upper left) and coral snake fang (lower left) and their corresponding cross sections. Photos from Plessis, Broeckhoven & le Roux, GigaScience vol 7, 2018
In general, viper fangs are very long and slender, with closed thinner walls that are more curved than coral snake fangs. The latter tend to be thicker, with the walls not completely fused — a characteristic of evolutionary trait shared by snakes related to coral snakes.
More intriguing are the differences in the angles and tips of the fangs. Sharp edges exist in all fangs. The viper has sharp edges along the top and undersurfaces, but only at the very tip. The coral fangs have small lateral ridges on each side of the fang (almost half way from the tip up) .
Mechanical stress tests using artificial metal punches mimicking snake fangs in varying shapes, length, angle and tip contours confirmed that the slender the fangs, the better the piercing. Not obvious, however, was that the sharpness of the fangs depends more on the tip angle and not its contour. A narrow fangs with blunt/rounded tip is a better performer than a wider fang with pointed tip. Ergo the fearful efficiency of the slender, curved and very sharp viper fans.
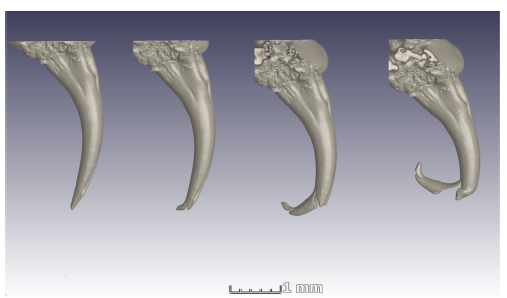
Fig. 7 is a series of microCT images showing what happened when the fangs were put under too much stress. To keep them sharp and ship-shape, fangs are replaced every two months.
Fig. 7 Micro CT of cobra fang. Illustration from same authors of Fig. 5
The following is the link to an excellent (and astounding) case history of a coral snake bite, an ordeal suffered by this Colombian physician https://doi.org/10.1016/j.rcae.2016.02.010
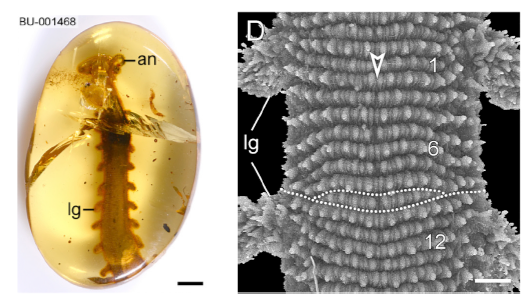
The first time I photographed this unusual slug/worm-like critter was during the construction phase of our Nectandra Cloud Forest Preserve in 2005. The 7.5 cm (3in) creature was slowly ambling around the wet algal carpet. It was brick red, with diamond shaped dorsal markings, purplish antennae, and a slightly flattened soft body, girdled with fine rows of “nubbins” on its skins (magnified view in lower photo B) and feet. It moved on two rows of 30 stubby, unarticulated, soft appendages. What was it? I was stymied. It had no exoskeleton, so couldn’t be a centipede or millipede. It couldn’t be a caterpillar because caterpillars have only 8 pairs of legs. It couldn’t be an annelid or slugs, because true worms or slugs have no legs.
Fig A (top) & B. Provisionally identified as Epiperipatus isthmicola , photographed at Nectandra, 2005
I soon learned that it was commonly known as a velvet worm. It belonged in its own phylum Onychophora, named for their clawed feet. Each stubby, retractable foot is a hollow structure covering several sets of sharp claws that are stacked as in Russian dolls. The outer set is shed successively during each molting. The claws are for positioning and traction.
Onychophorans are a group of enigmatic organisms believed to be rare. True enough, I had to wait a full decade to see my second photo subject. In between those years, I only knew it by its common name. Few live specimens had been studied globally at the time and their family tree was being hotly debated among biologists.
Almost another decade went by. We still know little of the onychophorans because they are shy and infrequently seen. They are mostly nocturnal, although the two velvet worms I photographed appeared both during the day. What little we do know hints of the importance of the velvet worm’s placement in evolution. Interest revived recently among a handful of evolutionary biologists (mainly in Europe and Costa Rica) to learn about the velvet worms phylogenetic relationships to their nearest relatives — the arthropods (insects, arachnids, crustaceans), which constitute 80% of modern earth’s fauna.
What do we know about the ancestry of velvet worms?
They are from a very ancient lineage, and are often called living fossils having survived the multiple mass extinctions since the Cambrian. The earliest known fossils were no less than 500 millions years old. Tiny (≤ 3 cm) ancestors of the velvet worms, remarkably well preserved for soft-tissue organisms, were found in the Canadian Burgess Shale (Figs. C-E) These Cambrian ancestors, informally grouped as lobopods (Greek for blunt feet) shared stubby legs with two other ancient groups — the ancestral tardigrades (water bears) and arthropods (insects, spiders, centi/millipedes, crustaceans).
Fig. C. Burgess shale. Claire H. from New York City, via Wikimedia Commons
Fig. D (upper) and E (lower). Lobopod Aysheaia pedunculata Walcott, a Cambrian onychophoran specimen collected in 1911 from the Burgess Shale. The specimen was photographed at two different light angles.
Fig. F Composite interpretive drawings of the specimen in Figs. D and E, by HB Whittington(1978). Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences Vol 284, No. 1000 Nov 16, 1978.
Anatomically, the onychophorans share many features with not just the tardigrades and arthropods, but also worms and nematodes. With no definitive identifying morphological traits, the phylogeny of the velvet worms languished for years, until the advent of DNA analysis and other non-destructive imaging technologies to study the 100 million yr old Burmese amber samples (Fig. G). We are finally beginning to glimpse into the lives of these curious organisms.
Fig. G Amber specimen from Myanmar, Burma. Left, Light microscopy. Right, Dorsal study of the same specimen by X-ray microtomography ( SRµRT). http://dx.doi.org/10.1016/j.cub.2016.07.023
Where do the onychophorans live
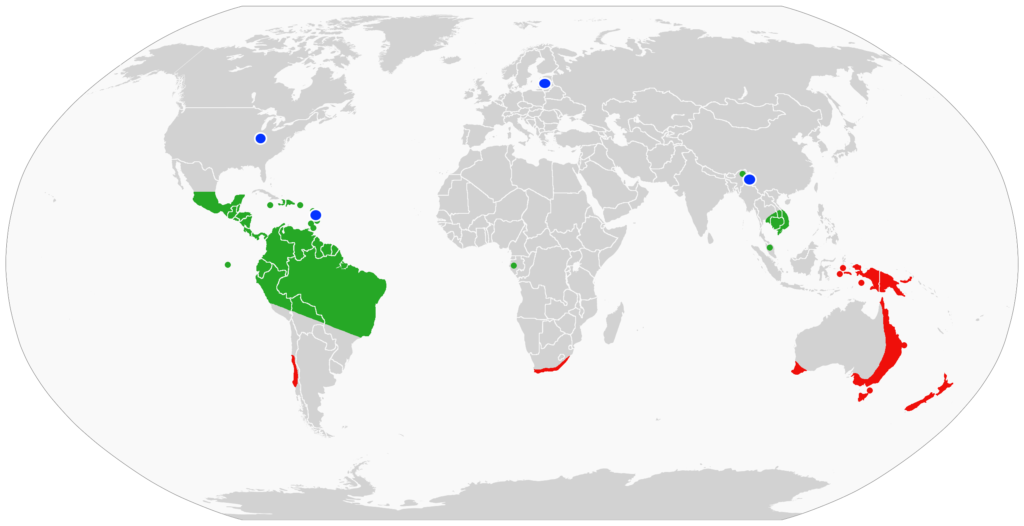
Without an exoskeleton, the terrestrial velvet worms are susceptible to desiccation. They are found in dark, damp, rotting plant litter or crevices in both the tropical and temperate wet forests. Their low density and infrequent sightings have hampered investigations into their biology, habitats, ecology and behavior. While we know about 200 species worldwide, most of the known natural history of the velvet worms comes from a period of interest between 1826 -1900. The more recent renewed attention and information indicate that there may be many more species yet to be discovered.
Fig H. Global distribution of onychophorans. Green and red areas represent two major subgroups. Blue dots are locations of fossils. From: Benutzer Achim Raschka via Wikipedia Commons.
The largest number, 74 described species, of velvet worms is in Australia. A sampling of the Australian onychophorans show an array of resplendent velvety dorsal cuticles (Fig. I).
Fig. I Australian Onychophorans. Original photographs by Jenny Norman, Noel Tait and Paul Sunnucks. Current biology Vol 21 No7.
Unusual biology of the Onychophorans
The biology of velvet worms is nothing short of extraordinary.
Mating — Onychophorans reproduce sexually (except for one known hermaphroditic species). Actual mating has only been observed for 2 Australian and 1 Costa Rican species, with different variations on the mating positions.
In the Costa Rican species, the male and female join their abdominal ends for the insemination through the female vagina — the direct approach.
In one Australian species, the male deposits a packet of sperms (spermatophore) on the female skin on her back or sides. The spermatophore covering and the female skin are digested by the female’s enzymes to allow the sperms to migrate through the female body walls, and eventually through the ovary walls to fertilize the eggs — the indirect approach.
In the other variation, the male uses a special, ornate head structure to press the spermatophore directly into the female vagina (Fig. J). There is no information how the spermatophore is positioned on the male’s head in the first place. The female supports the male’s head with her hind claws while he positions the spermatophore. They remain joined for 15-30 min and could move as a joined pair, during which time the sperms are released into her vagina — the head-to-tail approach.
Fig J. Australia onconyphorans mating: the male inserts the spermatophore vaginally with its head. Right: detail of the male’s head with its yellow spermatophore. Drawing by J. Monge-Najera based on photographs by Tait and Norman (2001). https://revistas.ucr.ac.cr/index.php/rbt/index
Thus far, there has been only one investigation into the fate of sperms after mating. Anatomically the female uterus is equipped with long-term sperm storage pouches containing sperms from multiple males, as well other accessory pouches containing decomposed sperms with mysterious ciliated funnels connecting the two.
Depending on the species, diverse reproduction of young is either by egg-laying or young-bearing. In the egg-laying species, females lay large yolk-filled eggs, while other species retain the eggs until they are ready to hatch. In the young bearing species, yolk-less eggs are nourished in the placenta. When the young are fully developed, the mother gives birth to her young similarly to placental mammals.
Feeding — They feed on small invertebrates and are voracious carnivores, hunt at dusk, subsisting on small invertebrates, worms and snails.
Hunting — The onychophorans most unique trait is their prey capturing technique. When feasible, they sneak up on the victim, poke it with its antennae to assess the prey’s size and risk. Alternatively, the velvet worm detects approaching prey through air movement. When ready to attack, it rears up its head, aims two large glands just under the antennae, then fires two gyrating jets of instant glue-like protein under high pressure. The fast acting, fast drying adhesive entangles the quarry within seconds. The velvet worm then spends many hours feasting afterward on the prey and consuming the nutritious slime. The latter is too nutritious to waste.
The following link is a short video showing how one physicist studies the fire-hosing technique in microscopic detail.
Nectandra Reserve’s location is well known among the local old timers as a garden of Eden — and just as snake infested. This reputation was especially well heeded by parents. Children were warned off and even forbidden from entering the place in the pre-Nectandra era. Several of these children are now among our garden staff.
As you may have read from my last blog, the snake skin pictured was found wrapped around the clothes dryer in our Costa Rican home. It necessitated a most inconvenient and unusual session of lint removal. That experience, coupled with the numerous snake sightings at the Nectandra Reserve, triggered a series of musings. In turn, they led me to an unexpected wealth of fascinating research information.
Given the number of snakes at Nectandra, finding abandoned skins is a common occurrence. Fast growing young snakes molt several times a year, while older snakes do it annually. The skins are found hanging off tree branches or on the ground. The photo below, of a longer than 2m (~6.5 ft ) venomous pit viper (Fer-de-lance or Bothrops asper), was collected by our staff and spread on the measuring board marked off with 5 cm (2in.) squares. On the same surface were also negative plaster-of-Paris casts of Baird’s tapir footprints and of two different, very large cats, later identified as a puma and a jaguar.
Snake skins are also disconcertingly common around my home near the reserve. Yet, they almost always induce my heart to skip a beat. I encounter them in the most unexpected places, such as on the handle of my propane tank, the fan of the clothes dryer, between the planks of lumber in my shop or in the middle of the garden gravel path. At first glance, these places shared no apparent common attributes. Aren’t snake supposed to be extra shy when they are molting? Another perplexing observation was that skins, if found intact, were completely turned inside out. This suggested that the snakes didn’t just shed their skins willy-nilly, but “crawled” out the narrow tube (through the mouth). The old skin and the new skin must also be attached at the tail until the inversion is complete. Intrigued, I searched for web videos of snakes in the process of molting. For a peep show of one caught doing the striptease, please click on the following link:
In other words, the snake needs a location with precise geometry and texture, a place where it can squeeze through easily but tight/rough enough to retain the old skin, such as branching tree forks, or if not available, a handle of a propane tank, a clothes dryer fan, a gravelly surface, or any suitable location at hand — ergo, the listlessness before molting trying to find a perfect spot. Incidentally, in the absence of suitable “shedding” structures, pet snakes can develop health problems associated with incomplete or imperfect shedding.
What happens during molting?
I described in my blog “Out of Its Skin” some of the reasons why certain organisms need to molt — mainly to make room for growing and elongating, for renewal of a protective skin subjected to continuous friction and to shed potential parasites.
A couple of weeks before shedding, snakes become restless, hide and stop eating. The old skin loses its luster. The eye shield (a transparent scale) clouds up (left photo) with a blueish tinge. This eye cap, known as the brille, is joined to the rest of the skin. The photo on the right shows the brille of a snake after ecdysis (molting) at Nectandra.
In their growing phase, the outer old skin and the inner new skin are contiguous. Unlike human skin, where one individual aging cell sloughs off at a time, snake epidermal cells form a single interconnected sheet, to be shed all at once. For shedding, the highly keratinized (analogous to our nails) old skin needs to separate from the more delicate inner layer. Just before separation, white blood cells (think pus) invade a temporary zone between the two layers, releasing enzymes to digest the adhering tissues to facilitate molting.
The Old Skin
Snake (live or dead) skin is dry to the touch, moderately flexible and scaly. Plainly, the scales are not like those of fishes. They cannot be scraped off as they are folds in the integument and are an integral part of the skin. Depending on the body location, the scales are of different size and shapes (photos A & B. ). The precise distribution and shapes of the scales are used to identify snakes.
A. Inside out snake skin molt. Dorsal head skin to the right of fold. Ventral head scales to the left. B. Dorsal scales (upper) above banded ventral scales (lower).
Scales
“Wait a second”, you may say, “but snakes are not colorless! Where are the snake patterns?”. Missing in the dead skin are the scale pigments that give the snake’s impressive coloration and camouflage. The pigmentation of the scales is critical, not just for appearance, but for the snake’s survival. The photos below show an example of the astounding ability of the W. African Gabon viper to disappear from view in their ecological niche. Snake pigments, composed of melanin, ranging from yellow to dark brown to black, are designed both for camouflage and temperature control. The darker the color, the better to absorb surrounding heat.
a. West Africa Gabon viper (Bitis rhinoceros) without background, b. on leafy substrate. c. Dorsal scale , d. Light microscopy of dorsal scale cross sections. Adapted from M Spinner et al. 2013, Scientific Reports.
While the dorsal scales determine the snakes appearance and radiation absorption, the ventral scales take most of the beating being always in contact with solid substrates. The keratinized scales not only are tough, but shaped to maximize gripping power when necessary and minimized friction for efficiency. In fact, ventral snake scales are not passive structures. They can be actively oriented to resist downward sliding, e.g. when climbing or going downhill (see photos below).
(A) A corn snake ascending a tree. (B-C) Scales are used to grip tree bark asperities. Snake scales at their (D) minimum and (E) maximum angles of attack. (F-G) A snake climbing an inclined surface. Sliding is prevented by emergency braking associated with lifting of the body. Adapted from PhD thesis of Hamidreza Marvi, 2013. Georgia Institute of Technology.
Now, we come to a key structure that enables the snakes’ undulating locomotion: its skeleton, which consists of the skull, vertebrae and ribs. Snake upper and lower jaws are not fused together. There is no sternum. Snakes have between 130-500 vertebrae. While each vertebra have limited freedom of rotation, there are enough of them to collectively allow for the snakes’s overall flexibility. Each of the hundreds of vertebrae is attached to a rib. The other end of the ribs, without a sternum, are attached to muscles and skin. Together with their unfused jaws, expandable rib cage and skin, snakes can swallow and ingest preys many times their own size — whole.
A snake skeleton in National Museum of Natural Sciences of Spain
Locomotion
In a moment of fantastic reflection, I imagine living in an exceedingly long, limbless body, straightjacketed by a shapeless tube. I ponder what it would take for me to slither, climb, swim or even glide in the air (as in the flying snakes ) — all the while constrained inside a body that is ~20 times longer than my girth. Snakes may have lost their limbs during evolution, but they certainly have made up the loss in astounding ways — feats beyond my human comprehension.
Moving snakes are hypnotic to watch. The serpentine locomotion is smooth, silent, fluid, swift yet unhurried (even when pursued). Let’s view a video clip of a coral snake (~2.0m) moving across a freshly dug construction site at Nectandra. Note the fast rhythmic undulation of the body, known technically as concertina movement. In the concertina mode, the body forms S-shaped undulation horizontally, but also coupled to shallow, vertical folding and unfolding movements using anchored contact points with the substrate to propel forward. In the video below, the entire length of the body appears to “flow” over the rough terrain without any apparent effort. The unhesitating, liquid-like movement belied the unevenness of the substrate, even on the steep incline. (I could attest to the terrain roughness because I did not do well on the same path). Imagine the frictional demands on the skin.
Coral snake, approximately 2.0 m long, slithering across construction site at Nectandra. Videographer Arturo Jarquin.
Not surprisingly, the versatility of snakes’ locomotion has engineers in awe and in eager pursuit to understand the forms and functions of the snake structure and skin. They are especially curious about the skin’s counteraction to unrelenting friction, as well as other anatomical features that allow the high degree of articulation. The engineers’ curiosity is not simply academic, but to inspire designs for industrial search-and-rescue robotics, surgical applications and even space exploration. Snakes, as it turn out, are among the most efficient, flexible and versatile locomotors, requiring the least space over a wide ranging substrates. Many of their cleverly designed robotic cousins hope to emulate the ability to squeeze through inaccessible crevices, building rubble, pipe lines, collapsed tunnels and mine, human blood vessels or organs etc.
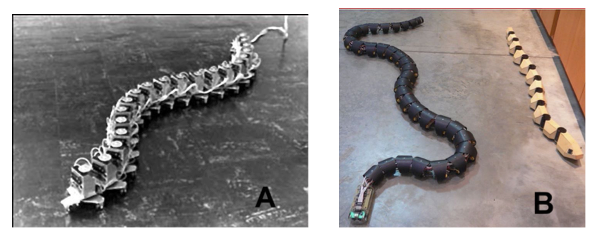
In trying to tease out the details of snakes locomotion, engineers have been building snake robots since 1972, to investigate and emulate the constituent components that influence the snake articulation and locomotion. There have been innumerable designs for snakebots, many with wheels, rollers, even legs. Some are powered by electricity, others propelled by pressured air and fluids etc. The experimenters learned rapidly that the effortless slithering, climbing, swimming of biological snakes belie the enormous complexity in their dynamic movements, body structures and surface components. The following photos are just a small sampling of robots among many.
A. Earliest robot, ACM iii, 1972. Shigeo Hirose of Tokyo Institute of Technology B. Robo-snakes, photo by Steve Jurvetson, 2005.
C. Robot Uncle Sam 2010, Carnegie Mellon University. D. Ladder climbing robot 2008. Tatsuya Takemori, Tokyo University & University of Electro-Communications
The robots photos in A, B are examples of snakebots (or robosnakes) that can slither on a smooth, flat surface but not climb. Robots in photos C and D can move, climb but not slither. There are innumerable others emulating the snake with varying degree of success, but all imperfectly compared to their biological versions.
I am including two very informative (and entertaining) videos of examples of man’s ingenuity. The first is Snakebot Engadget, a newer generation of Uncle Sam (robot C above), with impressive abilities already used in disasters search-and-rescue missions.
The second video (equally worthwhile) is of a large series of snake robots demonstrated at the 2021 Expo held at the Tokyo Institute of Technology, to sample the wide ranging ideas that have been tried.
Recently, our clothes drier developed an overheating problem. It was simple to diagnose the culprit as a malfunctioning blower fan, but it was a rude shock to discover a 3 feet snake skin, strung across the fan blades, with the head end to the right and out of view in the photo. I was impressed by the intact and clean appearance of the skin, a sign that its owner used the dead fan as the anchor for shedding after the fan died. That was not all. Imagine our surprise to find an additional snake corpse — one similar in size to the first — curled tightly in a ball plastered by caked up lint underneath the drum. This one died, with innards and vertebrae intact, perhaps of heat stroke? (I will come back to snakes molting in a later blog).
These types of inconvenient events are frequent but normal part of living in the tropics. I have learned to take them in stride and even turn them into sparks to light my curiosity fuse. This particular one started me thinking about how differently we humans shed skin.
The outermost layer of our body, the skin, is often referred to as the largest human organ, weighing about 8 lbs (3.6 kg) and constitutes the second largest surface area (intestinal surface is first). It is a protective barrier to prevent water loss and to shield against harmful sun radiation, heat and cold, chemicals and pathogens. But more than a passive barrier, it produces vitamin D, exudes antibacterial substances, and insulates. It is packed with fat cells (fuel, insulation and padding), Langerhans cells (immune sensors), nerves, sensory receptors and blood vessels. Depending on the location, it also has hair follicles and various glands (sweat, oil etc.). Our brain and body depend on it for cues of the outside world, to regulate body temperature, blood flow and our first line of immune defense. We cannot live without it.
Humans grow from two cells (one each from mom and dad) to around 30 trillion cells on average at maturity. To accommodate the increase in body bulk, not to mention the microbiome that we carry around, we shed our skin continuously, seamlessly, effortlessly, one cell at a time, 500 million (~ 0.002%) cells daily for the duration of our life.
This continuous skin renewal is contingent on the presence of the outermost layer, the epidermis. At the base of the epidermis are continuously growing cells. These upward migrating cells eventually die and slough off. Without an intact epidermis, as when injured or diseased, normal human skin regeneration does not occur and results in scarring.
In contrast, growth development in most invertebrates proceeds in multiple, disruptive steps, requiring sharp changes in form and function. Each stage of growth (known as an instar) is necessary to allow for the animal’s enlarging body. Each instar is protected by a skin, and at the final stages, by hard exoskeletons. These transitions take time and energy, during which the animal is vulnerable to attack and predation.
My Jan. 18, 2020 blog showed some of the changes during development of the moth Arsenura batesii, whereby the skin sloughed during the transitions between the caterpillar to pupal stage. The current blog looks at the skin shedding of the cicada.
Cicada
For many visitors to Nectandra at certain time of the year, it is difficult to ignore the synchronized ear-piercing “love song” of the male cicadas (females are silent), the world’s loudest insect, to attract mates. This sound is generated by two oscillating (300-400 times per minute) membranes called tymbals, one on each side of the abdomen just underneath the wings. After mating, the cicada eggs — laid usually on tree branches or trunks — mature, hatch and become nymphs. The emerging nymphs drop to the ground where most of the development takes place. For most species, the nymphs spend 2-5 years underground but some stay as long as 17 years. Not all cicadas have synchronized emergence. Many species have annual cycles. Below are photos of three cicadas species (yet to be identified, out of 45 in Costa Rica) found at Nectandra.
The last subterrestrial nymphal stage eventually emerges, climbs on a suitable tree trunk with camouflage potential, then proceeds to change into its last coat for its mating performance. The next photo captures the perfect out-of-body moment of the cicada emerging from its transparent skin. The different reflected colors off the old and new body from the camera flash give the scene the surreal effect. During ecdysis (skin shedding), the hard exoskeleton split neatly in the back, from which the next stage emerges. Note the body size difference between the transparent cast and the newly emerged cicada.
Often, hundreds of cicada abandoned casts can be seen still attached to trunks of trees.
Ecdysis is a risky process for the animal. For example, the pressure from the pooling hemolymph (fluid analogous to blood) that is needed to split the backside of the exoskeleton (photo below) sometime fails to do the job, causing the emerging cicada to get stuck. Or, parts of the body are injured
during the exit. Or the cuticle of the new body could not harden properly (a three day job) — a condition vulnerable to predation. Ecdysis is very energy intensive. In case of the cicada, the old exoskeleton is abandonned, unlike that of amphibians. The anolis below, in contrast, ate its proteinaceous skin upon shedding.
So, what are the advantages for the multiple cicada development stages? For a start, the soft-bodied nymphs get protection and humidity from being underground where they can attach to tree roots to suck the sap. The variable time underground avoids food competition between the young and adults, and other cicada species. Synchronized emergence from soil facilitates the hunt for mates. Thus, safety, food and sex optimized — just differently from us.
How much can one see (or miss) in a blink of an eye ?
This topic arose when my spouse David and I were on a garden walk around our Costa Rican residence. After weeks of misty and windy days typical of the cloud forest mountainside, the sun burst through the clouds, in and out, on this particular morning. The glorious weather drew us out of the house. We were standing on a slope where the photo below was taken, surrounded by moderately tall trees. Unlike temperate gardens often with a profusion of colors, our tropical patch is solid green, from the ground up to the treetops. It is an assisted regenerating forest on former ranch land. At that time of the year, there was not a single speck of color among the green foliage.
While luxuriating in the soft sun and breeze, we saw a bright orangish-yellow, midsize Sulphur butterfly flying horizontally in straight line at altitude of 10-12 ft high (3m), from right to left in the photo, aligned with a strong breeze and heading toward us. It reached somewhere at the center of photograph, and poof it vanished. At that moment, nothing unusual was moving within view except for that single butterfly.
In retrospect, both of us were synchronously tracking the only bright thing around and were seeing the exact same thing. Or rather, not seeing, the same thing as neither of us saw what happened. We must have both blinked at the precise moment when the butterfly disappeared. Not understanding what occurred, we kept our puzzlement private and continued with our walk.
My own mind was slow to process what happened. Did I really “see” it disappear or did it drop out of the sky? But there was no butterfly on the ground. Was Mother Nature practicing her magic trick? What happened? I scanned the ground again, but unable to find a quick explanation, I puzzled on.
From the corner of my eyes, I saw David loitering around the vicinity (near the small tree on the right edge of the garden photo) where the butterfly disappeared. Five minutes later, I heard him call “Evelyne, come quick. You have to see this”. Camera in hand, I headed for the spot. He pointed to a small tree. I followed his finger tip but saw only shiny, green leaves. No flowers, no buds, totally unexceptional. “What should I be seeing?” I queried. “Step back and look again”. Three steps backward and sure enough, the significance popped into view. Not all the foliage was green! One lone leaf was turning color.
I homed in on the yellowed leaf and there was our Apricot Sulphur (Phoebis argante ) clinging to it (next photo below). The breeze was blowing very hard. It was struggling to stay vertical and barely hanging on.
When it detected our presence, the butterfly naturally sought refuge. As far as our eyes could see, that dying leaf was the only speck of yellow. In 1/3 of a second (duration of a human blink), it spotted the sole yellow leaf, flew to it, landed, and hid in plain sight. Its evasive tactic would have succeeded spectacularly if David wasn’t so piqued.
As further proof that it was in the “hide” mode, it stayed on the leaf during my entire photo session. Because of the wind, I fiddled for 5-10 minutes trying to get a sharp close-up photo, staying within 2 ft. My heart went out to the butterfly. It fought hard and hung on valiantly during that whole time. Anyone who has ever tried to photograph butterflies can attest that this was abnormal behavior, as they are exceedingly skittish and camera shy animals. It finally flew off when we retreated at least 15 ft .
I learned recently that visual processing involves one fourth of our human brain. Relative to the rest of our body, it is an astounding proportion to support the small eyes. The butterfly brain, in comparison, is pinhead size. Keeping this in mind, let’s work out roughly what it would take to achieve the feat, in human terms, and compare score of each trait using symbols to mean equal (=), human better than butterfly (>), or less than butterfly (<) respectively.
High flying speed
The incredible speed the sulphur butterflies zip around with a tail wind at our location has always fascinated me. I clocked them at 8-10 mph under similar breezy conditions. Taking into consideration the relative body sizes, this would roughly correspond to 2X the running speed of a champion athlete.
Number of eyes
As we know, we human have two eyes of one type designated as simple eye (as in single lens). Butterflies, on the other hand, have 2 simple eyes (hidden on top of the head in photo below, at the base of the antennae) and 2 large compound eyes (each with arrays of 12,000 individual tiny photoreceptors known as ommatidia). The butterfly simple eyes sense light intensity, movement, and orientation whereas the thousands of compound eyes can sense shapes but mainly colors (see below discussion on Colors). Contrary to popular belief, butterflies see only a single image, not thousands with the compound eyes. Each eye contributes a “pixel” to the whole image.
Close up of Papilio spp. Photo credit: Nitin Jawale
Biologically, vision refers to the process whereby the eyes convert incident light into electric signals that are eventually processed by the brain. Vision allows animals to be aware of their physical surroundings. In butterflies, it is especially dominant, more so than the olfactory or auditory senses. Vision in general and color vision in specific are critical for survival. The butterfly depends on visual cues to feed on their preferred flowers, find mate, reproduce (where to lay eggs) and to migrate.
We can now break down and compare the separate components involved in human and butterfly vision.
Visual distance and depth of field
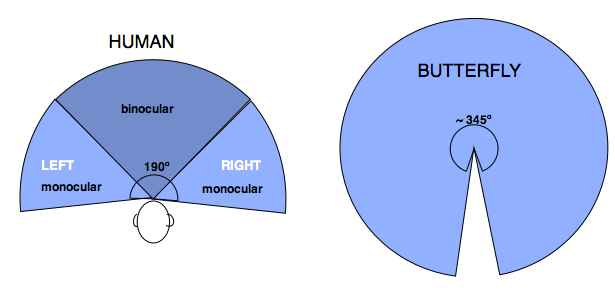
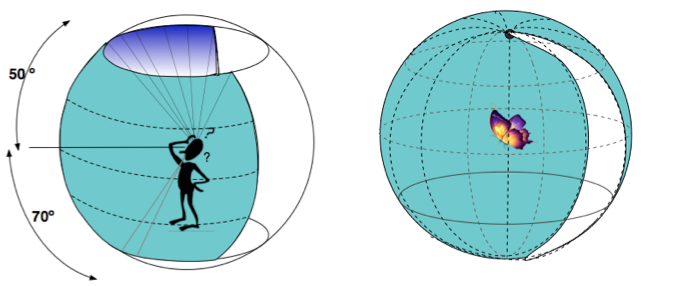
The position and structure of the two human eyes allow for binocular vision with depth perception, whereas butterfly’s eyes are capable of monocular vision. In distance, man can see much further, 3 mi vs. 12 ft for butterfly. For the purpose of the discussion of our disappearing butterfly, let’s keep to a distance of 12 ft for comparison.
Peripheral Vision
Peripheral vision is what we can see out of the corner of our eyes. The boundaries of butterfly peripheral vision are substantially greater than man’s. Our vision is confined to the space immediately in front, shown below in the cross section of the peripheral field. We cannot see behind our shoulders or above our head. The butterfly numerous eyes can see everything from front to back except for a ~ 15º arc behind its head.
In 3-dimensional space, this difference between human and butterfly is even more pronounced. Human can see well ahead within 120º vertical arc. Butterfly can see in front, above, below, and behind except for the blind arc — amounting to 93% of the visual sphere. This was gleaned from a study of the Orange sulphur butterfly and may not be true for others species. In comparison, human can see less than 30% of the visual sphere. (The latter is my rough calculation).
Rough respresentation of human and butterfly visual sphere
Visual Acuity
Acuity allows us to resolve small details with precision (think Snellen eye chart at the optometrist office). The next illustration shows the difference between human and butterfly. Among studied animals, human visual acuity is among the highest. Butterfly rates somewhere in the middle.
Photo credits: Ron Rutowski ; Modified from illustration nsf:Multimedia Gallery
Color and Light Sensing
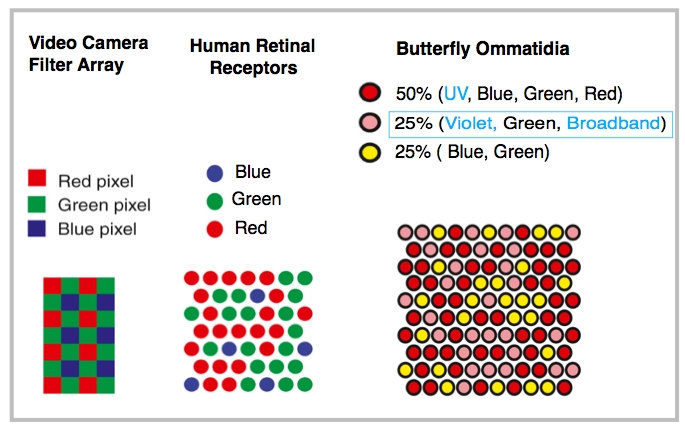
Extreme vision is the butterfly’s ace in the hole. What it lacks in brain cells, it makes up in the number and types of different photocells. Butterfly visual range may be the widest among animals. Their photoreceptors are vastly more numerous, and more significantly, they are more varied (chart) in types and in sensitivities to a wider spectral range. For example, eyes of the studied Japanese yellow swallowtail can detect blue, green and red (as in humans) but also violet, shorter (UV) wavelength, longer (broadband) wavelength and polarized light.
The next illustration shows the relative arrangement and distribution of the human receptors (cones) and butterfly ommatidia (packets of 13-15 bundled receptors) in comparison to the digital video camera on the left.
Adapted from Almut Kelber, Current Biology 26:R900-902, 2016
Butterfly’s more varied and more complex photoreceptors support a variety of functions. For example, UV light enhances their visual differentiation of flowers as food sources. Polarized and broadband light are important in the recognition of potential mates and in signal communication among a world of other butterflies. They also depend on the wider spectral range to search and recognize plant sources, especially for tropical butterflies in a highly biodiverse, dark and dense forest environment.
The following butterflies are photographed under visible and UV light.
Photo credit: Adrian Davies
The next two sets of photos illustrate the appearance of flowers and of butterflies in visible light, UV and simulated vision to fellow butterflies. We cannot, of course, know what the butterflies “see”, but we are beginning to learn what they can detect and what they respond to.
Photo credits: Klaus Schmitt
Photo credits: Klaus Schmitt
The above are just a handful of the traits that affect butterfly visual perceptions, studied on a small number of butterflies out of the 17,000 species total.
If you have kept count, the score so far is butterfly 4, human 2.
And that, dear readers, is why we missed the show.
There is no escaping the mosquitoes, even in the cool and misty cloud forest. Certainly in comparison to lower elevation, the density of mosquitoes at the Nectandra Reserve is low, and thankfully, does not include either Aedes aegypti or albopictus, the fearsome vectors of several human pathogens (yellow fever, dengue, chikungunya and Zika etc). But disease carriers or no, mosquito bites are infernally itchy.
Both male and female mosquitoes feed on sugary liquids (nectar, aphid honeydew, fruit juices etc), but only female mosquitoes suck blood to obtain protein for their eggs. We have all experienced that pin prick sensation when she inserts her long, slender and stiff proboscis through our skin. Below are photos (Fig 1) of the proboscis before and after removal of the opaque outer cuticle. The mosquito was being prepared for a synchrotron X-ray imaging technique to study the dynamics of blood suction in vivo. The bared proboscis in Fig. 1B is actually a bundle of six stylets — one pair to hold on to the victim’s skin, another pair with saw teeth to cut through it, the fifth is a probe for the victim’s blood vessels and the last, a straw-like suction tube which is the actual food canal.
BH Kim, HK Kim & SJ Lee. J Expt Biol 214, 2011
With the proboscis in position, she injects 1 µL of a potent mix of salivary anophelin proteins (sometime comingled with pathogens) to numb the pain of the bite and thin the host’s blood for easier suctioning. Incidentally, the itch receptors are only found in the top two layers of human skin. Surgical removal of those skin layers removes the capacity to itch! That explains why we do not feel the itch in the muscles.
To pump blood the mosquito does not use her heart, an organ dedicated to her circulatory system. Instead, blood suction is accomplished through a different pump mechanism located in her tiny head. It consists of two interconnected but separate pumps .
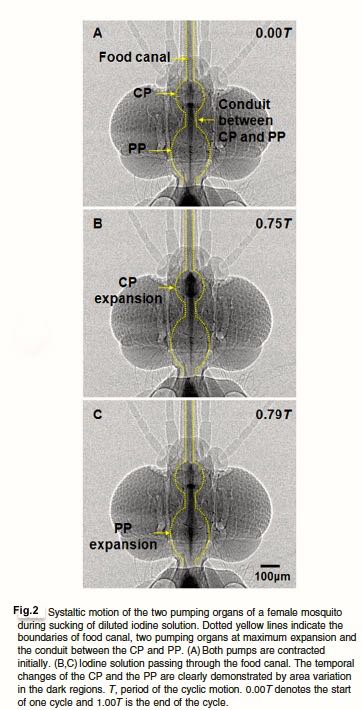
Below is a series of micrographs showing the travel of an opaque iodine solution (to simulate blood) through the food canal and through the first smaller pump (CP) and second larger pump (PP) in the Fig 2.
BH Kim, HK Kim & SJ Lee. J Expt Biol 214, 2011
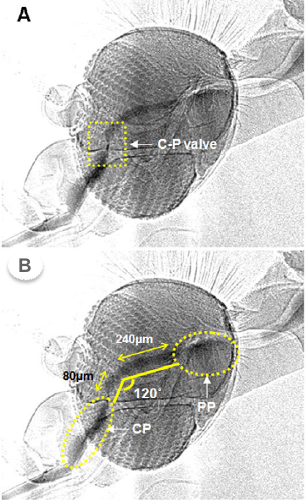
Why two pumps and not just one? Turns out the anatomy of the mosquito head requires the blood to course through a sharp 120º bend and a check valve (C-P, Fig. 3 ) to reach the gut. The first smaller pump CP is just efficient enough to get the viscous blood through the bend and valve. The large pump PP, already inflated and ready, then propels it trough the digestive system . The two pumps in series are superbly optimized for the task.
Fig. 3 BH Kim, HK Kim & SJ Lee. J Expt Biol 214, 2011
The impetus for this blog was triggered by a startling observation on a nightwalk at Nectandra not too long ago. My hiking companion spotted a Pug-nosed Anole lizard (Anolis capito) perched on a branch. She took the photo below of this fairly common diurnal species in our forest. The 5 in. lizard (torso only) almost always rests aligned with the tree branch or trunk, with its legs retracted close to its body. Its green speckled marking is often hard to spot during the day because of its excellent camouflage.
I, too, lined up to take a shot while my companion aimed her flash light to provide me with extra light. I zoomed in with my macro-lens. To my utter astonishment, the lizard was not alone! I spotted four tiny mosquitoes on the lizard, two on the upper torso and two near the tail .
Magnified 5X on the computer screen, the photo was even more telling. The mosquitoes must have been feasting for quite some time. They all appeared to be fully engorged but obviously unsatisfied and still feeding. Their stomachs are filled with hemolymph and quite extended. Full engorgement takes about 3 -10 minutes. Meanwhile, the immobile lizard looked oblivious, or perhaps just resigned.
There are more than 3000 species of mosquitoes worldwide. They feed on a wide range of hosts — mammals, birds, reptiles, amphibians and fish. Their feeding patterns can be broadly classified into: a) specialists that feed exclusively on warm-blooded animals or cold-blooded animals but not both, b) generalists that feed on both, and c) opportunists that feed on varying available hosts depending on geography and climate.
Take the one infamous mosquito genus Aedes on which we have the most information. Two species within the genus, aegypti, once found only in African tropics, and albopictus from southeast Asia, have now spread to all five continents, even to temperate regions. They have different ecologic niches, biting habits, and host preferences. Aegypti feed mostly on humans, but sometimes cattle, pigs, cat, rat and chicken. Albopictus, on the other hand, feed exclusively on humans. Because of their preference in biting humans, they are in effect transferring the pathogens from an infected human to a susceptible human. Together, they contribute to half million human deaths annually from the four diseases mentioned above.
Things are quite a bit more complicated in Eastern equine encephalomyelitis, a rare mosquito-borne viral infection in horses and humans that is often fatal due to brain inflammation. Here several genera of mosquito are involved as “bridge” vectors. They transfer the pathogen from the sources that are mainly birds, but also reptiles and amphibians — to susceptible humans and horses. There are no human-to-human transmissions.
From just the little we know of mosquitoes as major vectors of human diseases, it is not hard to extrapolate the power of the flying hypodermic needles to mix pathogens and match hosts, probably including our Pug-nose lizard.
Aside: I couldn’t help feel a sympathetic itch looking at my photo of the lizard, and was curious to know if amphibians and reptiles itch from mosquito bites. My literature search yielded only one Australian observation describing bitten frogs in the wild, mostly on eyelids, do not get irritated by the mosquitoes bites. However, they do end up with inflamed swelling on the eyelids.
Gustavo and his horde of relatives invaded Nectandra Reserve completely undetected. What snagged our attention was the defoliation of one tall Wercklea insignis tree (related to the Hibiscus family) with the dinner-plate size leaves. Under subterfuge, Gustavo and clan were eating the foliage nightly, but vanished by daylight. At first, the consumption was insignificant but gradually increased exponentially. Over the next two weeks, the culprits had become veritable feeding machines. The leaves were mowed at an astounding rate each night, but now the thieves were leaving an impressive amount of droppings on the ground by morning. Caterpillars! Judging from the size and appearance of the frass (insect poop), we knew they were from large moths, but which moth? There are some 5000 known ones in Costa Rica. Flightless, how could caterpillars disappear so completely each night? Easy. Hide in plain sight.
Once fingered, our caterpillars were spotted even in daylight. I felt utterly foolish and blind for missing them in the first place. There were many dozens of caterpillars of the same species, large as cigars, all aligned and with the exact coloration of the tree trunk! Immobile after their nocturnal feeding frenzy high up near the canopy, their striking tentacles were like beacons once we knew what to look for. Intrigued, my co-worker decided to adopt three selected specimens from the dozens. We wanted to follow the development to the end of their metamorphosis to identify them. Moths are identified morphologically from the adult and not the caterpillar stage. After transferring them to a large plastic container, the caterpillars were presented with fresh Wercklea leaves each night. Then we watched.
Naming the study subjects is a double-edged sword — a no-no among scientists for fear the anthropomorphism would cloud the observers’ judgment — but we couldn’t help ourselves. The handsomest of the three, we named Gustavo. He was already 10 cm in length (4in) not including his impressive head gear when we spotted him. He had 4 pink-black-white banded head tentacles on his head and one black-tipped pink (absolutely charming) curlicue on the tail segment. The photo of Gustavo below was taken on the first day (D1) of our study. Judging by its large size, Gustavo was probably already nearly full-grown.
We also chose two other fellow caterpillars to hedge against mortality during captivity. Jorge (right upper in photo below) was chosen because he had one distinctive broken left-head tentacle and Freddy was smallest in size. We needed a way to tell them apart.
By Day 11, Gustavo grew only slightly, but his spots were lighter in color, larger, sharper and more noticeable.
Left. Gustavo on D11 Right upper. Jorge with a broken tentacle Right lower. Freddy
Four days later, D15, Gustavo shed its cuticle (skin) with the tentacles attached. He now looked very different. Its retractable head was beginning to enlarge, his spots fused and the triangular green patches faded. He was also acquiring a layer of fine facial hair. Jorge and Freddy in the meantime had not yet molted (photos below). The typical moth or butterfly sheds its skin 4-5 times. The stage in between each molt is known as an instar. The fourth or fifth instar is usually the final instar before it turns into a pupa if it is a moth, or chrysalis if it is a butterfly.
Things were now moving faster. By D18, Gustavo and Freddy were no longer eating. Both were shrinking in girth and length. Only Jorge still had his tentacles, was still feeding and growing.
The photo of Gustavo on D27 revealed the classic body of caterpillars (next photo). First the head on the left is connected to the thin 3-segmented thorax, behind which is the thicker 10-segmented abdomen. The first three thoracic segments have three pairs of pointy, true legs, one pair per segment. These true legs help position the head while rasping the leaf during feeding. Thoracic segments 3,4,5,6,10 are equipped with pairs of prolegs (not jointed as the true legs). The muscular prolegs are equipped with clawed hooks for grasping. Anyone who has tried to dislodge caterpillars from their food plants knows how tenaciously the subjects can cling to the leaves.
Caterpillars have six single-lens eyes on each lower side of the head. These eyes can sense light intensity but do not form images. For respiration, they have a system of internal tubes and valves that end in spiracles (valves) on each side of the thoracic segment for gas exchanges (black dots in photo above visible on each segment). Inter-caterpillar communication is by deposition and emission of chemical trails. The caterpillars’ sensitivity to plant aromatics is incredibly efficient. I once witnessed a coordinated U-turn by a pack of 3 dozens caterpillars within seconds after I introduced fresh food plant within 15 cm (6 in ) of the lead caterpillar.
The next photograph (D27) below show the variation among their siblings not in captivity, still clinging to the tree. The narrow range in size indicated they were likely all hatched from eggs laid about the same time. Note the individual coloration and markings that allowed them to blend in the tree trunk —another example of the art of concealment mentioned in my previous blog. After all, they are sitting “ducks” at this stage, completely vulnerable to predation from birds, bats and parasitoid insects.
D 27. Wild cousins of Gustavo on the tree branches
The formation of the pupae was pretty mysterious. Gustavo stopped feeding altogether shortly after D28. By D47, it has shrunk and hardened, but still with recognizable segments and prolegs. The photo below shows all three in the prepupa stage: Jorge was still caterpillar in form, whereas Freddy has started to pupate. Gustavo is still shrinking. By this point, the caterpillars had consumed enough nutrients and conserved enough water to last them through a complex series of transformation events to reach the terminal moth stage, most of which are not yet fully known.
The next image shows all three on D67. Gustavo was clearly a pupa showing hints of internal structure underneath the translucent exoskeleton, Freddy in the intermediate and Jorge in the semi-pupa stage.
At this point, not knowing the identity of our subjects meant we had to guess how to maintain the three pupae alive. Moth pupae are of two types: 1) naked pupae, usually found among leaves or soil or 2) in silk or spun cocoon. Since ours were of the first type, we put them in individual pots of loose soil and leaves. We then waited, and waited, and waited…
A full eight months later in October, resplendent Gustavo slowly and carefully emerged from the 5 cm pupa. Finally, we know what it is — a moth in the Saturniidae family, Arsenurabatesii .
Gustavo emerged
Gustavo had an impressive 13 cm (5in) wing span. Its body and wings were elegantly and spectacularly furry. The hair on moths are not truly hair, but modified scales that are also found on butterflies. The scales serve multiple functions, for thermoregulation, sex appeal, camouflage, and as expendable body structures during attacks. Nocturnal predators such as bats have been known to end up with mouthful of fur but not the prey.
This is where the decision to name our study subjects comes home to roost. Gustavo should have been named Gustava, a female, based on the thread-like antennae. The roving male saturniids have complex, feathered antennae, better to sniff out the mostly stationary, pheromones advertising females. Saturniids moths do not eat and have reduced mouth-parts. They only need to stay alive a couple of weeks — long enough to find a mate, and lay their eggs.
What is the benefit of metamorphosis? The most obvious is that the caterpillar and the adult moth occupy totally different niches. No competition between the old and the young. The two use different food sources, are totally different in forms, locomotions and phase duration. They are not subject to the same risks and vulnerabilities.
Our little exercise revealed only the stages between the last instar and pupa. We were not privy to the mysterious events behind the exoskeleton of pupae before moth emergence. The totally cloaked metamorphic events are just beginning to be unraveled. Thanks to micro-CT scanning adopted for the purpose of imaging interior events, we soon will have much more information of this astounding process.
It took us nearly a year’s observation to document one single caterpillar-moth pair. Imagine the colossal work of Dan Janzen (U of Pennsylvania) on 9000 moths in the last three decades in Costa Rica, not just their biology (photo below), but also DNA barcoding for final identification .
Dan Janzen’s investigations on moth metamorphosis, using plastic bags housing for caterpillars with their food sources. The photo was taken in one of Dr.Janzen’s several growing sheds in Guanacaste, Costa Rica, on my visit in 2011.
Brrrrr…. It’s 52 °F in our cloud forest this morning. Not cold by northern-latitudes standard, but downright frigid for the tropics, especially for the two dozen species of Nectandra hummingbirds (of some 50 in Costa Rica).
Imagine the challenge in thermoregulation for an average size (4 in) hummingbird such as the Rufous-tailed hummer pictured below with a body weighing as little as a single grape (0.18 oz).
As most birds, it must maintain body temperatures between 100-108 °F to stay active. For these birds, foraging for food means to be able to keep heartbeats around 1200 beats/min to sustain wingbeats of 50-70/sec to fly at average speed of 34 mph. This flight ability allows the hummingbirds to hover about the flowers for nectar during feeding, and even to fly backward — unique acrobatic locomotion among avifauna. (Human: 60-100 heartbeats/min and jogging speed of 4-6 mph )
Anatomically, the hummingbirds have the largest relative heart to body size. The rufous-tailed’s heart constitutes 2.4% of its body weight (compared to ~0.5% of average human ). Its need for oxygen during flight translates into higher red blood cell density of 6.59 million/ml (~5 million/ml for humans).
To stay fit, hummingbirds must eat and drink a lot — more than half its body weight each day in
nectar and twice in water, the equivalent of some 1000 flowers/day (favorite
examples among many blossoms at Nectandra in photos below).
Top left to right: Columnea microphylla, Guzmania spp, Sobralia virginalis Bottom left to right: Elleanthus robustus, Justicia aurea, Besleria notabilis
Critical protein, fat and mineral supplements come from small insects, fruit flies, spiders, larvae, insect eggs, etc. (examples on Sobralia orchid, photo top right). These food items are extra-critical during egg laying and feeding of average clutch of two young birds (when the mother birds themselves do not eat).
In addition to the extra caloric need during motherhood, some hummers must store fat (about 0.1 oz), enough for 24-26 hr of long flights (longer than the 22 hr limit for Boeing 777 non-refueling flight). For example, the Ruby-Throated hummingbirds migrate across the 500 miles Gulf of Mexico. They can clock at 25 mph or 650 miles maximum, enough to cross the Gulf of Mexico non-stop with energy to spare. Rufous hummingbirds can migrate 1865 miles (with stops) between Mexico and Alaska/Canada.
The high metabolic rate creates interesting physiologic challenges. For one, the high caloric intake must be metabolized, channeled and expended fast enough not to overheat. As result, the hummingbirds have vastly efficient sugar digestion, increased tissue gas transport and matched respiration rate. To give an idea of what it takes to achieve the same high level of metabolism, an average human would have to consume 100 lb. of pure glucose per day. The corresponding human metabolism, however, would generate lethal body temperature equivalent to ~752 °F .
The tiny-bodied hummingbirds must work at marginal and exquisitely tuned energy balance. For example, to counteract the risk of overheating, hummingbirds do not have down feathers, even for those in the temperate-zone during winters. Instead, they rather conserve energy over long period, as during sleep, by going into a short hibernation state known as torpor — a lowering of body temperature to a steady minimum of around 60-70 °F, dropping 60-90% of its metabolic rate and >30% water loss in this torpor state.
Hummingbirds, in sum, are superbirds. Their rapidly adjusting metabolic rates, the highest of any animals, allow them to withstand great ambient temperature swings. They may be the smallest but have the largest heart, the greatest appetite and the sweetest tongues. They can fly the fastest and longest, besting the newest and longest range jet plane without refueling. They can hover and fly backward like no other, birds or machines.
Most of all, hummingbirds are undeniably among the most beautiful of birds with their flashy, iridescent plumage. Their feathers are wonders to behold — a topic worthy of another conversation. Please join me on a eye opening chin-to-tail hummingbird feather inspection in the next blog.